Bütün canlı hücreler zarın iki yüzeyi arasında bir voltaj ya da zar potansiyeline sahiptir.
Yani plazma zarı elektriksel olarak kutuplaşmış yani polarize olmuştur.
Bunun anlamı zarın bir yüzeyinin diğerine göre daha negatif olduğudur.
Zarın iki yüzeyi arasındaki bu potansiyel farka zar potansiyeli adı verilir.
Dinlenme durumundaki bir nöron hücresi -70 mili voltluk bir zar potansiyeline sahiptir.
Burada işaretin negatif olması hücre içinin dışına göre negatif yüklü olduğunu belirtir.

Zar potansiyeli, Na+ ve K+ iyonlarının hücre tarafından kontrollü şekilde hücre içi ve dışına taşınmasıyla sağlanmaktadır.
Bunun yanında hücre içinde bulunan anyonlar da ( proteinler, aminoasitler, sülfat, fosfat ve diğer negatif yüklüler ) zar potansiyelinin oluşmasında etkilidir.

İyonlar yağda çözünmedikleri için fosfolipit tabakasını difüzyonla geçemezler.
İyonların zardan geçişi ya zardaki taşıyıcı proteinlerce pompalanmaları ya da iyon kanalları boyunca pasif olarak hareket etmeleridir.
İyon kanalları özgül iyonlar için seçicidir.
Zarın potasyum iyonlarına karşı geçirgenliği, sodyum iyonlarına geçirgenliğinden daha fazladır. Çünkü zar üzerindeki K+ kanallarının miktarı daha fazladır.
Hücre içindeki anyonların çoğunluğu büyük moleküllü proteinler olduğundan zardan geçemez.
Dolayısıyla hücre içinde büyük bir negatif yük havuzu oluşur. K+ iyonları bu yük havuzu tarafından hücre içine çekilirken, ortaya çıkan konsantrasyon farkı nedeniyle de hücre dışına çıkmak ister.
O halde hücreye K+ giriş çıkışı bu iki kuvvetin etkisinde gerçekleşecektir.
Bu duruma hücre tarafından müdahale edilmezse hücre içi ve dışı için Na+ ve K+ iyonlarının konsantrasyon farkları kaybolacaktır. Bu durumda zar potansiyeli oluşmaz.
Zar potansiyelinin oluşturulması, hücredeki Na – K pompası tarafından sağlanır.
Bu pompa sürekli olarak Na+ iyonlarını hücre dışına, K+ iyonlarını ise hücre içine taşıyarak -70 milivoltluk zar potansiyelinin oluşmasını sağlar.

Tüm hücreler zar potansiyeline sahip olmasına rağmen, aralarında sinir ve kas hücrelerinin de bulunduğu belirli hücre tipleri zar potansiyellerinde büyük değişimler oluşturabilme yeteneğine sahiptirler.
Bu hücrelere topluca uyarılabilen hücreler denir.
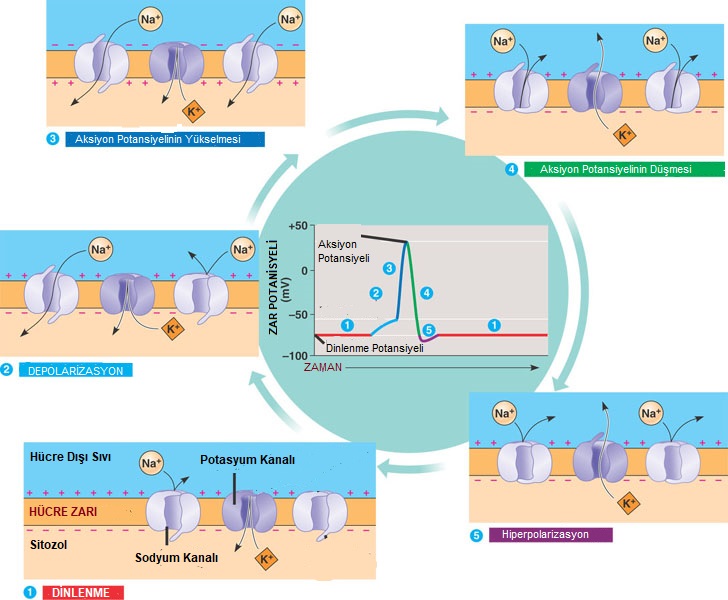
Dinlenme durumundaki bir uyarılabilen hücrenin zar potansiyeline dinlenme potansiyeli adı verilir.
Uyarılabilen hücrelerde zar potansiyelinin değişime uğratılması bir uyarana tepki olarak açılan ya da kapanan kapılı iyon kanalları sayesinde gerçekleştirilir.
Bu kanallar sürekli açık olan kapısız Na+ ve K+ kanallarından farklı yapıda kanallardır.
Kapılı iyon kanalları sadece tek bir tip uyarana cevap olarak açılır ya da kapanır.
Örneğin kimyasal kapılı iyon kanalları sinaps noktalarından salınan nörotransmitter maddelerin etkisiyle açılıp kapanırken, voltaj kapılı iyon kanallarının açılıp kapanması zar potansiyelindeki değişimler sonucu uyarılır.
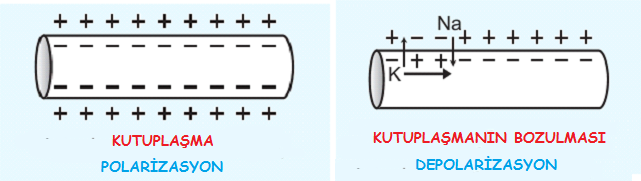
Sinir hücresi dinlenme halinde iken hücre dışı pozitif, hücre içi ise negatif yüklüdür.
Hücrenin bu durumu Na – K pompası tarafından sağlanır ve polarizasyon adını alır.
İç ve dış çevredeki uyarılar, sinir hücresinde impuls ( uyartı ) adı verilen elektriksel ve kimyasal değişiklikleri başlatır.
İmpuls oluşabilmesi için, uyarı şiddetinin eşik değeri aşması gerekir.
Sinir hücresinde impuls oluşumunu sağlayan en düşük uyarı şiddetine eşik değer ya da eşik şiddet denir.
Sinir hücresi bu değerin altındaki uyarılara tepki vermez.
Eşik şiddet ve üzerindeki uyarılara ise aynı şekilde tepki verir. Bu durum ya hep – ya hiç prensibi olarak adlandırılır.
Sinir hücresi aktivite gösterdiğinde yani impuls iletimi yaptığında zarın Na+ iyonlarına geçirgenliği yaklaşık 500 kat artar ve Na+ iyonları büyük bir hızla içeri girer.
Bu durumda polarizasyon bozulur ve potansiyel farkı +40 milivolta çıkar.
Bu duruma depolarizasyon denir.
Sinapslardan salgılanan nörotransmitter maddeler kapılı iyon kanalları üzerine etki ederek impulsun diğer sinir hücresine geçmesini ya da durdurulmasını sağlar.
Eğer impuls sonraki sinir hücresine geçecekse kapılı Na+ kanalları açılır ve Na+ iyonları hızla içeri dolar. Depolarizasyon durumu oluşturulur ve impuls iletilir.
Bunun aksine eğer sinaps durdurucu bir sinaps ise bu kez nörotransmitterler sonraki nöronda kapılı K+ kanallarının açılmasını sağlayarak potasyumun hızla dışarı çıkmasını dolayısıyla zar potansiyel farkının daha da büyümesini sağlayarak ( hiperpolarizasyon oluşturarak ) impulsun geçişini durdurur.
Sinir hücresi üzerinden impuls geçtikten sonra zar potansiyeli dinlenme durumuna tekrar dönüştürülür.
Bu durum ise yeniden polarize olma anlamında repolarizasyon olarak adlandırılır.

Uyarılabilir bir hücrenin zar potansiyelindeki hızlı değişme aksiyon potansiyeli olarak adlandırılır ve bu değişiklik impuls oluşumunu tetikler.
Depolarizasyon durumunda Na+ kanalları açıldığı için Na+ iyonları hızla hücre içine dolar.
Ancak belli bir süre sonra bu giriş durdurulur. Na+ girişinin durdurulmasından hemen önce K iyonları dışarı çıkmaya başlar.
Hücreye giren Na+ kadar dışarı K+ iyonu atılır. Ancak K+ kanalları yavaş kapandığı için zar dinlenme durumuna dönmeden önce bir miktar hiperpolarize olur.
Daha sonra Na – K pompasının etkinliği ile zar dinlenme potansiyeline tekrar kavuşturulur.

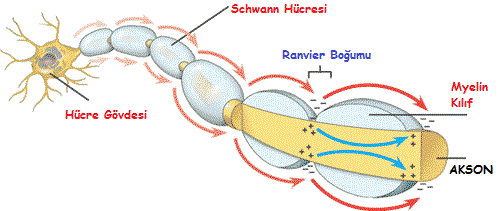
Sodyum iyonlarının hücre içine girmesi ile oluşan ilk aksiyon potansiyeli zarın bir sonraki bölgesinde de başka bir aksiyon potansiyeli oluşturur.
Aksiyon potansiyeli oluştuktan sonra akson boyunca ard arda devam eder. Yani impulsun iletimi aksiyon potansiyelinin ardı ardına tekrarlanmasıyla olmaktadır. ( Domino taşlarının devrilmesi gibi )
Aksiyon potansiyelinin akson boyunca tekrarlanma hızını akson çapı ve aksonun myelin kılıf taşıyıp taşımaması etkiler.
Akson çapı ne kadar büyükse iletim hızı o kadar yüksek olur.
Çünkü kalın bir aksonda belirli bir zaman dilimi içinde daha çok iyon akar ve sonraki bölgede daha çabuk aksiyon potansiyeli oluşturulur.
Omurgalılarda, aksiyon potansiyelinin tekrarlanma hızını artıran myelin kılıf evrimleşmiştir.
Myelinli aksonlarda iyon akımı sadece Ranvier boğumlarında olur.
Bu sebeple myelinli aksonlarda görülen iletime atlamalı iletim denir.
Myelin kılıf impuls iletim hızını yaklaşık olarak 10 kat artırmaktadır.
Myelin kılıf taşımayan aksonlarda impuls iletim hızı 12 m/sn iken myelinli aksonlarda bu hız 120 m/sn kadardır.
Bir nöronda iletilirken, elektriksel değişiklikler yanında kimyasal değişiklikler de olur.
İmpuls iletilirken O2 ve glikoz harcanması, bunun yanı sıra CO2 oluşup sıcaklığın artması kimyasal değişikliklerdir.
Uyarının eşik şiddette veya daha fazla olması impulsun hızını ve şiddetini değiştirmez.
Ya hep ya hiç yasasına göre eşik şiddetin üzerindeki uyarılar aynı şiddette impuls oluşumuna sebep oluyor ve bu impulslar aynı şekilde taşınıyorsa uyarının şiddeti nasıl algılanır sorusunun cevabı frekansta gizlidir.
Reseptöre ulaşan uyarının şiddeti ne kadar fazla ise nöronda daha sık aralıklarla impuls oluşturulur.
Duyu merkezleri gelen impulsun sıklığına bağlı olarak uyarının şiddetini belirler.
Örneğin sıcak bir cisme dokunduğumuzda oluşturulan impuls sayısı, ılık bir cisme dokunduğumuzda oluşturulandan fazladır.

Sinir sistemine gelen uyarının çeşidinin belirlenmesi ise uyarıyı alan reseptörün çeşidine ve impulsu taşıyan yola bağlıdır.
Örneğin ışığın fotoreseptörlerde oluşturduğu uyartı, beyindeki görme merkezine taşınır ve görüntü olarak algılanır.
Bir sinir ya da kas hücresi tek başına ya hep ya hiç prensibine uyar.
Ancak çok sayıda sinir hücresinden oluşmuş bir sinir kordonu veya çok sayıda kas hücresinden oluşmuş bir kas demeti bu prensibe uymaz.
Çünkü her sinir hücresinin uyarılması için gereken eşik değer bir diğerinden farklıdır.
Düşük şiddetteki uyarı öncelikle kolay uyarılabilen sinir tellerini veya kas tellerini uyarır.
Uyarının şiddeti arttıkça uyarılan sinir teli sayısı artacağından daha kuvvetli cevap verilir.
Belli bir değerden sonra ise verilen cevap değişmez.
PDF DOSYASI İÇİN
![]()